
— Аннотация.
Данная книга является продолжением моей ранее опубликованной «Математические подходы к оценке Эволюционного потенциала систем». Эта книга читателю предлагается посмотреть на самые разные вопросы с точки зрения Эволюционного потенциала. Наибольшее внимание уделено вопросу возникновения (создания) Искуственного разума (ИР). Хотя в данном направление происходит бурное развитие, пока остаётся без ответа вопрос: в какой момент времени компьютеры смогут стать носителями Искусственного Разума? Попытке ответа на данный вопрос посвящена часть этой книги. Технологии и способы возникновения ИР выходят за рамки этой книги. Не так уж и важно: возникнет ли он случайно в процессе усложнения систем или это будет результатом планомерных действий по его созданию.
Как я уже упомянул, в качестве рабочего инструмента для анализа будет использоваться описанный мною ранее Эволюционный потенциал и подходы к расчёту его величины.
— Ограничения.
Дадим рабочее определение Искусственного Разума.
Под Искусственным Разумом понимаются свойства системы к взаимодействию с окружающим миром через его непрерывное познание, имеющее свойства отображения мира в виде своих собственных образов, манипуляции с этими образами, предсказания поведения и свойств элементов окружающего мира, способности создавать абстрактные и новые, принципиально не похожие на уже имеющиеся, модели систем и элементов мира, а также манипуляции с ними в целях закрепления и расширения достигнутых результатов. Также должны проявляться как постепенные, так и скачкообразные усложнения способностей к дальнейшему познанию.
Скажу сразу, что поскольку нас в основном будет интересовать уровень сложности системы, позволяющий проявиться ИР, мы сосредоточимся в области его оценки и в меньшей степени уделим внимание подходам в достижении данного уровня. Пути создания вычислительных систем, способных к эволюционированию в меняющейся среде, и, как результат, возникновение в них ИР затрагивать не будут. Не будет и прогноза даты очередного «апокалипсиса» в результате «Революции ИР».
— Методика и последовательность действий.
В качестве некой опоры, на которую можно опереться в оценках сложности системы — носителя ИР, предлагается рассмотреть особенности человеческого (биологического) разума. В таком подходе нет ничего нового. Уже стало традицией сравнивать мозг и компьютеры с точки зрения обработки информации. Новым будет здесь оценка Эволюционного потенциала (ЭП) человеческого разума. Далее можно прогнозировать, что возникновению ИР будет соответствовать похожие значения эволюционного потенциала у технических систем. Я сознательно не пишу «компьютерных», поскольку не факт, что ожидаемому значению будет соответствовать компьютер в том виде, к которому мы привыкли. Но даже при всей внешней простоте такого подхода имеется множество подводных камней потенциально способных не дать довести задуманное до конца.
Затем, имея на руках требуемую величину эволюционного потенциала технической системы, далее можно будет попробовать рассмотреть способы, которыми её можно достичь.
— Эволюционный потенциал разума.
Чтобы оценить ЭП человеческого разума, логично вначале обратиться к таблице в предыдущей книге, по которой делается оценка ЭП. В таблице мы находим значение 1, соответствующее строке «Формирование и достижение цели на уровне разума». Это уже достаточно близко к тому, что нам нужно.
Однако не все так просто. Эта величина слишком маленькая для человека. Человеческий мозг реализует намного больший набор решаемых задач и функций. Поэтому, прежде чем перейти к рассмотрению структур мозга и расчётам его ЭП, требуется отобрать те усложнения, которых достаточно для появления разума, отбросив то, что имеется у неразумных существ. И в этом кроется большая ошибка, поскольку любой вид разума не является статической системой и не существует сам в себе независимо от среды. Разумным делает систему ряд особенностей вплетения её в окружающий мир. Но данный вопрос уводит нас в плоскость философии и не хотелось бы в этом утонуть. Поэтому примем значение ЭП неразумных животных равное 1 (использовалось в расчётах ранее) и возьмём его как базовое на котором далее строится ИР.
Сложив эти два значения по ранее изложенным методикам, получим значение равное 2-м. Т.е. грубая оценка ЭП ИР находится в районе 2-х.
Вернёмся к мозгу. Можно определить величину Эволюционного потенциала, которому соответствует мозг как орган, но с потенциалами его периферических отделов типа глаза, уши и пр. возникают трудности, поскольку они не существуют независимо, а являются частью иерархической системы.
С этой точки зрения использовать значения ЭП из ранее сформированных таблиц по теме Эволюционного потенциала неудобно, поскольку они ориентированы на оценки потенциала с опорой на связи, а не сложность структур типа мозга животного. Лучше сформировать под нашу задачу новую таблицу.
Сначала используем существующее. Будем рассуждать так:
1- величина ЭП разума человека появилась как результат развития мозга высших организмов, мозг которых реализовал все основные функции выживания и размножения. Т.е. он был уже достаточно развит, чтобы строить картину мира и её прогноз на основе сенсорных каналов получения информации и моторики на базе встроенной в обратную связь прогноза. Определив ЭП животного равным 1, этот потенциал также можно разделить на две части, одна связана с организмом как таковым, вторая будет соответствовать мозгу животного.
Почему в данном случае мы рассматриваем весь мозг, а не его отделы, которые в основном отвечают за высшую деятельность? Дело в том, что при рассмотрении мозга с точки зрения обработки информации, она уже начинается на периферии (пример сетчатки) и далее может ветвиться и обрабатываться в разных других отделах. К сожалению, нет достаточных для наших расчётов данных о разнице между устройством и обработкой информации мозга животного и человека в плане сложности, количества структур и связей.
Поскольку у нас нет таблицы, на основе которой бы рассчитывался ЭП живых организмов, сделаем дополнительные расчёты.
Количество клеток человека 1014.
ЭП одной простой клетки равно 0,01.
ЭП человеческого организма как многоклеточного равно (1+14) *0,01 = 0,15. Замечу, что нижнее пороговое значение для простых многоклеточных организмов определено в районе 0,1, а для интеллектуальных многоклеточных организмов (животных) в районе 1. Этот промежуток заполняется путём усложнения многоклеточного организма через формирование различных подсистем. С одной стороны они связаны друг с другом, с другой функционально «ортогональны» в плане независимости. Например, «зрение» и «моторику» (передвижение) можно считать ортогональными до некого иерархического уровня мозговой деятельности, даже с учётом переплетений за счёт различных низкоуровневых связей, реализации инстинктивных функций и т. п.
Количеству нейронов 86*109 человеческого мозга соответствует нижнее значение ЭП 0,12. Кора больших полушарий человека содержит 16*109 нейронов и имеет нижнее значение ЭП 0,11.
ЭП интеллектуального организма равно 1. Рассматривая разницу между насекомыми и животными, мы видим некий условный порог 250 тыс. нейронов у муравья и дальнейший рост у более разумных видов. Затем у животных происходит заметный рост количества нейронов и появление еще одной структуры в виде коры головного мозга. Если устойчивым признаком интеллекта у животных считать наличие коры головного мозга, то у грызунов количество клеток мозга 6*107 (ЭП 0,088), а коры их головного мозга 107 (ЭП 0,08). При этом количество всех клеток мыши примерно 3*1010.
ЭП мышиного организма равно 0,11. Отметим разницу ЭП мозга и организма всего в 0,03 единиц ЭП, что не удивительно, так как в наших расчётах не учитывается сложность мозга, увеличивающая ЭП мыши до величины близкой к 1.
Также в организации жизнедеятельности и работе нейронов участвуют и другие типы клеток (глиальные). Т.е. система обладает большей сложностью, чем просто однородные одноклеточные соединения. Оценки соотношения их количества к нейронам заметно разнятся во времени и в разных источниках. Согласно наиболее свежим данным, её величину можно принять равной 2,3 от количества нейронов. Т.е. вместо нейрона мы должны использовать подсистему из нейрона и 2,3 глиальных клеток. Но мы не можем в расчётах просто пропорционально увеличить количество клеток, поскольку хотя глиальные клетки и участвуют в деятельности нейронов, однако имеют более прозаичные предназначения. Возьмём это пока на заметку, а требуемый перерасчёт сделаем намного позже.
Каждый из нейронов имеет от 103 до 104 связей. На данный момент времени хотелось бы иметь готовую методику их оценки, но пока непонятно как оценить сложность связей, имеющих не просто электро-химическую природу, а участвующих в обработке поступающей информации и одновременно формирующих эти структуры.
Примем в качестве рабочей следующую гипотезу: поскольку состояние нейрона меняется с учётом всех поступающих через синапсы «данных» и текущего состояния самого нейрона, а также то, что этот результат распространяется через связи нейрона на значительное количество других нейронов, совокупность соединённых между собой нейронов будут образовывать единую систему, использующую выходящие наружу связи своих нейронов как собственные.
Межклеточным взаимодействиям мы присвоили значение ЭП равное 10—2, соответственно ЭП нейрона получает за счёт ЭП связей дополнительный потенциал порядка 0,03..0,04. Это значительная величина относительно простой клетки, и вполне допустимая для многоклеточной системы имеющей ЭП более 0,1, но все же недостаточная, чтобы преодолеть границу ЭП 0,1.
Выделим наиболее очевидных «кандидатов», увеличивающих Эволюционный потенциал:
— уровень сложности клетки типа нейрона на самом деле заметно больше простой клетки, т.е. 0,1 и более;
— организация соединений нейронов образовывает большое число сложных структур (подсистем).
Для определения ЭП нейронов учтём множество функций, осуществляемых ими совместно с синапсами:
— способность менять пространственные конфигурации,
— участие в совместной деятельности с другими нейронами и клетками,
— самоорганизацию.
Применим подход, использованный ранее при определении базовых величин ЭП различных систем. Требуется систематизировать довольно большой промежуток усложнений от клетки до интеллектуального многоклеточного организма. Сформируем таблицу составляющих (базовых элементов) и соответствующих им связей для многоклеточных подсистем и организмов. В дальнейшем их можно уточнять по мере роста понимания темы и полученных результатов. В связи с большим диапазоном изменения сложности выделим в отдельную таблицу клетки (1) и группы клеток (2).
Далее возможны два подхода. В первом случае можно соединять клетки во все более сложные структуры и вычислять их эволюционный потенциал, стараясь учитывать его возможную скрытую часть. Во втором можно поступить также как и раньше, выделив промежуточную ступень усложнения клеточного сообщества, тем самым уменьшив величину ступени от клетки к многоклеточному организму. В нашем случае эта величина находится в районе 0,1 и отвечает она за простые многоклеточные организмы существующие как самостоятельные единицы, способные к жизни и размножению на уровне инстинктов (примитивное поведение) в среде с источниками питания и обеспечивающие совместный метаболизм, питание, защиту и размножении всех клеток внутри этого организма.
— Потенциал клетки.
Таблица 1. Составляющие Эволюционного потенциала клетки входящей в группу клеток.
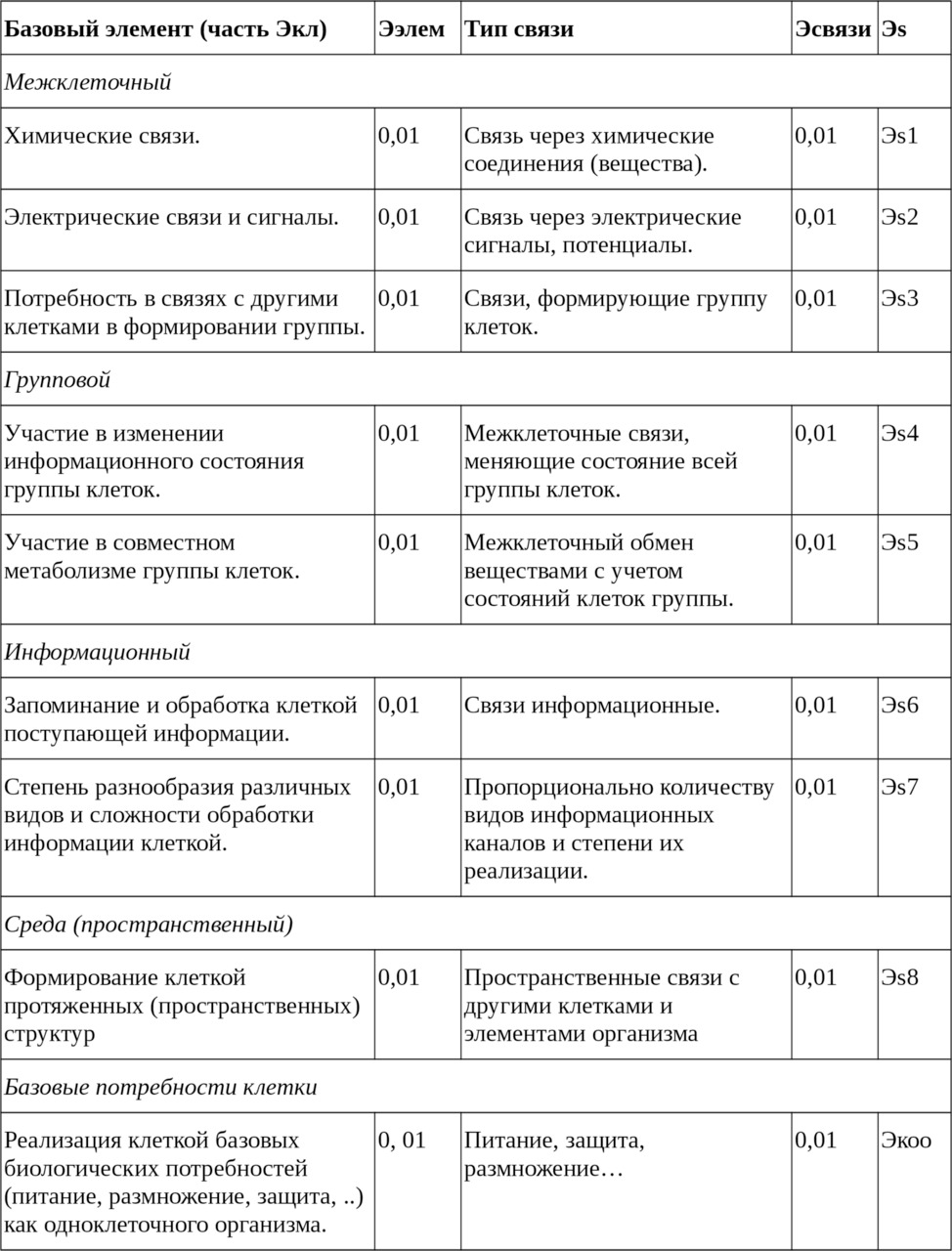
Из таблицы получаем достижимое значение ЭП сложной клетки, способной образовывать сложные сообщества (со сложным поведением) клеток, равное 0,09. За счёт наличия большого количества разных связей оно может достигать 0,1 и выше.
— Потенциал группы клеток.
Примем ЭП нейрона равным 0,1 (без глиальных клеток). В самом примитивном приближении человеческому мозгу, как набору связанных между собой нейронов, будет соответствовать ЭП=1,2. Кора больших полушарий человека будет иметь ЭП=1,1.
Чтобы понять, что простого увеличения количества нейронов мозга недостаточно для возникновения разума даже при максимальном количестве связей между ними, возьмём для примера слона. У него 257 млрд. нейронов в мозге и примерно 693 млрд. глиальных клеток. Даже если они образуют по 10 тыс. связей, все равно получим ЭП равным (1+12) *0,1 = 1,3 без учёта связей и максимально теоретически достижимое увеличение ЭП за счёт связей до 2,6. Что больше ЭП организма, но заметно меньше ЭП интеллектуального организма.
Чтобы количество клеток и связей перешло в «качество» можно организовать нейроны в группы, связанные друг с другом прямыми и обратными связями так, чтобы они совместно меняли свои состояния. В этом случае простой набор из нейронных подсистем изменяется качественным образом, образуя систему, которую можно рассматривать в первом приближении как гипотетическую многоуровневую (необязательно иерархическую) с точки зрения потоков информации.
Возникает предположение о том, что «разбивка» нейронов мозга на ряд областей, в сою очередь состоящих из нескольких слоёв, в которых клетки сгруппированы в локальные однотипные группы, а также слоёв, объединяющих эти различные области, создаст достаточную сложность, и мы сможем получить требуемую величину роста ЭП.
Определим те нейронные связи, которыми нейроны внутри одной группы связаны с нейронами других групп, как межгрупповые связи, поскольку группа нейронов через них реагирует на внешние сигналы как единая система и что влияние этих связей одинаковое. Полагаем, что нейрон меняет состояние, ориентируясь на удалённость и т. п. связи, а не на её принадлежность и что происходящие изменения распространяются на всю группу. Неопределённостью, связанной со степенью влияния этих связей на состояние самой группы, пока пренебрежём. Также считаем, что влияние всех связей одинаковое.
Для дальнейших расчётов определим вклад составляющей подсистему единицы (нейронов или группы нейронов) через значение Vед= W/Nед, если W <Nед или равным 1, если W> =Nед, где
W — количество выходящих нейронных связей из единицы, а Nед — количество единиц в подсистеме.
Рассмотрим примеры:
1. Расчёт увеличения ЭП нейрона с учётом связей через «свёртку» с ЭП других нейронов.
Рассмотрим однородную систему из N нейронов с величиной Эволюционного потенциала Эe и W связями с величиной Эсв.
Пусть n =1…N — номер нейрона, s = 1…W — связи нейрона с другими нейронами. W> 1.
Тогда каждый нейрон вносит вклад в образующуюся систему равный Vn= W/N.
Эсис = (1+log (N*Vn)) *Эew = (1+logW) *Эew.
Где Эew учитывает наличие связей и рост потенциала нейрона Эe за счёт связей.
Для случая Эсв <<Эe, используем упрощённую формулу:
Эew = Эe+ (1+logW) *Эсв.
Эсис = (1+logW) * (Эe+ (1+logW) *Эсв) = Эе* (1+logW) + Эсв* (1+logW) 2.
Например, для N=1010, W=104, Эe=0,1, Эсв=0,01, получаем Эew =0,15,
Эсис =0,75.
2. Расчёт ЭП большого числа параллельно расположенных групп нейронов из нескольких клеток (в виде столбиков), объединяемых верхними и нижними плоскими слоями.
Усложним систему группировку её элементов в однотипные подсистемы (группы), сделав разбивку системы на М групп с количеством нейронов в каждой из них Y> 1.
Если мы будем использовать значение Эew таким, как рассчитали раньше в 1 примере, то получится, что мы будем потом повторно учитывать потенциал связей, но уже как связи между группами, что не корректно. Поэтому Эew группы нейронов рассчитывается с учётом только внутригрупповых связей. Их немного и можно это значение взять максимально возможным. Т.е. каждый связан с каждым.
Тогда для группы можно записать:
Эew = Эe+ (1+logY) *Эсв.
Продолжая пример выше и учитывая, что Y = 10 и Эew =0,12
Эгр = (1+log Y) *Эew = 0,24.
Поскольку группа в совокупности задействует «во вне» все связи своих нейронов, за исключением тех связей, что окажутся внутри группы, при связях внутри группы «каждого с каждым» число межгрупповых связей, равное W-Wгр, уменьшится незначительно для W>> Wгр.
Используем значение Vгр= W/Nгр — вклад каждого нейрона для межгрупповых связей.
Продолжая пример, имеем Nгр = N/Y=109
Эсис = (1+log (Nгр*Vгр)) *Эгр = (1+logW) *Эгр = 1,2.
Отметим заметный рост Эволюционного потенциала системы по сравнению с первым вариантом.
3. Расчёт ЭП нескольких параллельно расположенных плоских слоёв толщиной в 1 нейрон.
Изменим группировку элементов системы, сгруппировав их в слои. Чем особенным характеризуется слой? Каждая клетка вместе с соседними не просто топологически расположена в локальной плоской области, но и количество её связей максимально в своём слое и меньше в других слоях.
Сама по себе топология расположения клеток нужна для физической реализации многослойной системы, мы же можем клетки слоя рассматривать как некую группу с описанной особенностью связей.
Пусть будет М слоёв с количеством нейронов в каждой из них N/М. Количество связей нейрона внутри слоя Y, наружу W- Y. Тогда для слоя можно записать:
Эew = Эe+ (1+logY)) *Эсв — потенциал нейрона.
Аналогично ранее написанному, вклад нейронов в слой равен Ve= Y/ (N/М), Y <N/М.
Эсл = (1+log ((N/М) *Y/ (N/М))) * Эew = (1+logY) * Эew — потенциал слоя.
(N/М) * (W-Y) — число межслойных связей, но поскольку фактически это связи нейронов, вклад потенциала их связей в потенциал слоя определяется обратной величиной их численности, т.е.
Wсл = W-Y.
Эслw = Эсл+ (1+logWсл) *Эсв — потенциал слоя с учётом связей (их достаточно много).
Vсл = (W-Y) /М — вклад слоя в образовании системы с учётом соотношения количества связей и элементов, если W-Y <М и Vсл=1 в противном (нашем) случае.
Эсис = (1+log М* Vсл) * Эслw.
Допустим в нашем примере М =10 слоев.
Y= 0,9*W =9*103.
Эew = 0,1+ (1+3,95) *0,01= 0,15, Эсл = (1+3,95) *0,15=0,74,
Эслw = 0,74+ (1+3) *0,01=0,78,
Эсис = (1+log 10) * 0,78=1,56.
Распределим связи нейрона поровну между внутренней и наружной частями слоя, т. е. Y=5*103.
Эew = 0,147, Эсл = 0,69, Эслw = 0,737, Эсис = 1,47.
Следовательно, увеличение соотношения количества связей внутреннего слоя относительно внешнего повышает потенциал системы, но этого все еще недостаточно.
4. Расчёт ЭП системы как комбинация вариантов 2 и 3, т.е. нескольких параллельно расположенных плоских слоёв, толщиной в несколько клеток.
Сложность здесь заключается в том, чтобы правильно учитывать доли вкладов подсистем, так как используются связи одних и тех же нейронов в разных подсистемах.
Допустим, имеются группы из 10 клеток, собранные в 10 слоёв (М). Для них можно записать: Y=10, Эew =0,12, Эгр =0,24.
Количество групп нейронов слоя Nгр =N/ (М*Y).
Вклад группы в слой. Число связей группы с другими группами внутри слоя равно Wгр = (W*Квн-Y) *Y, вклад Vгр =Wгр/ (N/ (М*Y)), если Wгр <Nгр.
Распределим связи нейронов поровну между внутренней и наружной частями слоя, т. е. Квн=0,5, Yгр=5*103—10, Wгр =5*104, Nгр =104/100 =102.
Поскольку Wгр> Nгр, Vгр =1.
Эсл = (1+logNгр) *Эгр = (1+2) *0,24= 0,76 — потенциал слоя.
Максимальное число межслойных (нейронных) связей без нарушения ортогональности для связей равно числу слоёв, т.е.10, поэтому за счёт добавки к потенциалу слоя потенциалов связей получим
Эслw = 0,76+ (1+1) *0,01=0,78,
Эсис = (1+log 10) * 0,78 =1,56.
Т.е. практически такое же значение что и в примере 3.
При всей ограниченности выше приведённых примеров можно сделать вывод о том, что «произвольная» разбивка на подсистемы хотя и даёт эффект в плане роста эволюционного потенциала, но наталкивается на проблему «параллельности» связей между подсистемами. Напомню, что сложности системы не растёт «сама по себе» при увеличении количества параллельных однотипных связей. Как бы не хотелось, но предложенные модели не обладают достаточным прогностическим эффектом в отношении конкретных способов реализации усложнения систем.
— Потенциал группы клеток многоклеточного организма
При определённых условиях расчёт ЭП системы, состоящей из «ортогональных друг другу в отношении роли и решаемых задач» подсистем, может осуществляться через суммирование их ЭП. Такое объединение реализовывается как единый результат взаимодействия (через цели и обратные связи) системы с окружающей средой. Это значит, что формируя в системе ортогональные по характеру подсистемы, осуществляющие разного рода и вида функции включая обработку информации, связанные между собой на разных уровнях и совместно реализующие основные целевые функции системы, можно многократно увеличить сложность и соответственно ЭП всей системы.
Таблица 2. Составляющие Эволюционного потенциала группы клеток как часть многоклеточного организма.
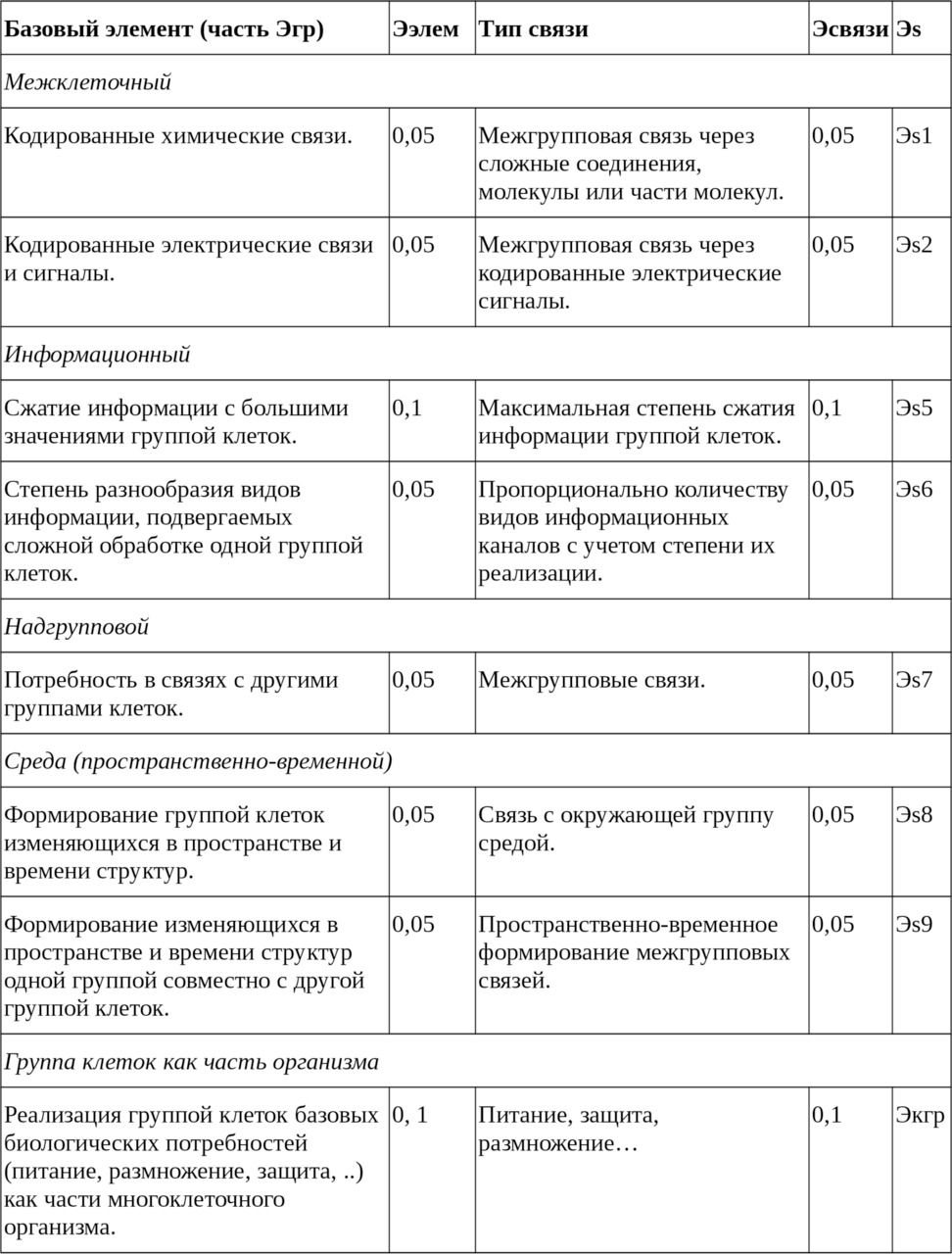
Как следствие выдвину следующую гипотезу: в искусственной системе существование ИР возможно с ЭП равным 1 и более при условии, что соответствующая ему подсистема будет сконструирована и создана с максимально мыслимо достижимой сложностью. Одновременно предполагается, что в своей деятельности она будет опираться на ряд других подсистем, обеспечивающих совокупный ЭП всей системы равным 2 и более.
В заключение этой главы можно сделать вывод, что появление ИР как побочный эффект повышения вычислительных мощностей и сложности программного обеспечения обычных компьютеров событие невероятное. Либо этого люди добьются целенаправленно, либо есть вероятность его появления в системах с ИИ, выполняющих проектирование и решение задач по комплексному управлению другими сложными интеллектуальными системами, по мере роста их уровня абстракции.
— Искусственный разум с точки зрения энтропии.
Потребление мозга оценивается в 20—25 Вт. Т.е. в секунду потребляется и рассеивается порядка 20 джоулей при температуре тела порядка 37 градусов С или 310 К.
Теперь сделаем оценку минимальных затрат на обработку бита информации при температуре тела 37гр. Если исходить из формулы Kb*T*ln2, где Kb коэффициент Больцмана, то она равна примерно 3*10—21 дж/с на 1 бит информации. Тогда мощность равная 20Вт теоретически позволяет обрабатывать 7*1021 бит/с.
Вернемся к мозгу. Если принять в расчетах условное количество нейронов 100 млрд, то в среднем на один нейрон со всеми его синапсами, спайками, глиальными клетками и т. д. придется 2*10—10 дж. Поскольку отсутствуют данные теоретического предела обработки информации элементами, имеющими реализацию в виде нейронов, поступим следующим образом. Оценим длительность одного «шага обработки» нейронами информации в 0,01сек. Соответствующее значение затрачиваемой энергии одним нейроном на один «шаг обработки» равно 2*10—12 дж. Тогда энергетический запас на переработку одного бита информации одним нейроном по сравнению с теоретическим пределом равен 1,5*109 на шаг. Замечу, это довольно большая величина.
Интуиция говорит, что даже при низкой, по сравнению теоретической эффективностью преобразования энергии в информацию и рассеянии соответствующей энтропии, получаем возможность обработки больших объемов информации. Например, при эффективности 1/1500 от теоретического предела можно обработать одним нейроном до 1 Мбит/шаг.
Хотя сравнение возможностей обработки информации мозгом и современными компьютерами делалось уже не раз, мы его в какой-то степени повторим для оценки ЭП компьютеров. Появившиеся нейроморфные процессоры не будем рассматривать по причине недостаточной информации в отношении больших систем на их основе. Рассмотрим классическую компьютерную архитектуру. Современный на данный момент времени двухпроцессорный сервер со 128 логическими ядрами в процессоре компании AMD потребляет (процессорами) мощность около 500 Вт. Оценим производительность этих двух процессоров величиной в 2,5*1012 операций/сек. Что достигается при возможности распараллеливания операций и предсказуемости команд ветвления. Получим затраты 500/2,5*1012= 2*10—10 Вт мощности на одну операцию. Т.е. если бы данный компьютер за одну операцию смог обрабатывать количество информации сопоставимое с нейроном, то умножив 2*10—10 Вт на количество нейронов 1011, получили бы значение 20 Вт как у мозга. Но поскольку для реализации одного «шага» компьютеру понадобится несравненно больше операций, его соответствующие энергозатраты при таких удельных значениях будут пропорционально больше.
Синтезируем некую упрощенную программную модель нейрона. Оценочно мы должны пошагово обработать у каждого нейрона синапсы в виде 10 тыс однословных параметров, каждый со своим весовым коэффициентом с учетом предыдущих состояний и состоянием самого нейрона. Не будем использовать такие сложные модели как антагонистические рецептивные поля. Объемом вычислений по обработке сложных состояний типа PSTH самого нейрона можно пренебречь на фоне обработки 10 тыс. параметров. Оценим размеры каждого из параметров и коэффициентов в 1 слово. Его размерность в байтах нам не принципиальна, поскольку универсальные процессоры оперируют стандартными словами достаточно большой длины.
На каждый параметр получится извлечение по адресу двух и более переменных, их сложения/перемножения и запись результатов в память, также мы должны добавить на каждую операцию еще сравнение адресов с границами, сравнение параметров с граничными значениями и команды ветвлений. Для обычного компьютера на это уйдет 20—30 и более шагов, поскольку распараллеливание здесь не эффективно из-за сильной последовательной связанности действий. Добавив временную обработку достаточно сложного поведения нейронов, количество выполняемых команд увеличиться еще больше. Допустим, удастся использовать локальное распараллеливание операций и остановиться на 20 операциях на синапс. Предположим, что удастся эффективно распараллелить обработку нейронов.
Тогда на всю обработку 10 тыс. синапсов и 100 млрд нейронов за 10 мс потребуется 104*20*1011 = 2*1016 операций. И в 1 сек нужно обрабатывать 2*1018 операций.
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.